![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)


Reproduction & dispersal
Vegetative Reproduction
Vegetative reproduction involves no mixing of genes from two parents. There is but one parent for the new individual. Many home gardeners are familiar with vegetative reproduction and practise it without necessarily knowing it by that name. A very common example is the growing of a new plant from a cutting. Similarly, many gardeners are familiar with the dividing of plants. A gardener who divides a plant to get two where there was previously one is again propagating vegetatively.
Bryophytes can reproduce vegetatively in various ways. In fact, it would almost be true to say that vegetative reproduction is the rule and not the exception. Vegetative reproduction is known from bryophytes where sexual reproduction has never been seen.
If the older parts of a branching or forking gametophyte die, the younger parts are left as separate individuals. The thalli of liverworts in the genus Riccia typically fork to produce Y-shaped growth forms. In due course the tips of each Y will fork in turn and you can see that beginning to happen in this photo ![]() . In Riccia the older parts die and decay, and you can see the early stages in this photo
. In Riccia the older parts die and decay, and you can see the early stages in this photo ![]() . In particular you can still see a hint of the Y shape on the left, but the thallus at the branching point is brown and dying. Soon you would have two, separate, strap-like thalli that would, in time, form new forks at their younger ends and so form new Y-shaped growth forms. This forking and dying process can lead to a rosette-like growth form for a Riccia colony, as shown in this photo.
. In particular you can still see a hint of the Y shape on the left, but the thallus at the branching point is brown and dying. Soon you would have two, separate, strap-like thalli that would, in time, form new forks at their younger ends and so form new Y-shaped growth forms. This forking and dying process can lead to a rosette-like growth form for a Riccia colony, as shown in this photo. ![]() .
.
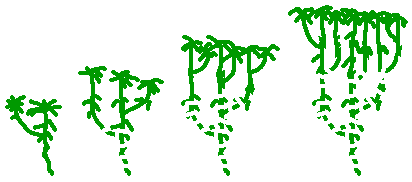
A similar process occurs in a number of mosses. Many mosses grow in compact colonies or mingled with other vegetation. In such circumstances, as the stems lengthen and branch the older parts may die and decay, creating separate plants where once there were branched plants. The accompanying diagram illustrates the process, showing just a moss and not the surrounds. Going from left to right a moss gametophyte branches. Both the main stem and the branches continue to grow. After some time the stem at and below the branching point dies. The dotted line indicates a former connection that has died. This now leaves two separate plants and the process continues. The final part of the diagram showing a number of separate plants of various ages. Sphagnum is an example of a genus in which this form of replication is common. This photo ![]() shows a view looking down on a small part of a Sphagnum bog and you can see a dense, self-supporting form of growth. What you see are the tufted heads of many Sphagnum stems. From the photograph you can't tell whether two neighbouring heads represent separate plants or two branches of a single plant. However, within this photo there will be Sphagnum plants at various stages of development shown in the earlier diagram.
shows a view looking down on a small part of a Sphagnum bog and you can see a dense, self-supporting form of growth. What you see are the tufted heads of many Sphagnum stems. From the photograph you can't tell whether two neighbouring heads represent separate plants or two branches of a single plant. However, within this photo there will be Sphagnum plants at various stages of development shown in the earlier diagram.
Most forms of vegetative reproduction involve some part of the gametophyte breaking off, being carried away by some means (e.g. wind, water, animal), landing somewhere else and then developing into a new plant. If a gametophyte fragment lands in a suitable habitat, it will grow into a new gametophyte. There are even fossils (dating from about 11,000 to 20,000 years ago) showing new plants growing out from moss gametophyte fragments![]() .
.
The bits that are capable of growing into new plants vary in form. They may simply be pieces that have been accidentally broken off. They may be ordinary parts of the plant that are pre-disposed to breakage. They may be tiny, specialized bud-like outgrowths that bear no resemblance to any "ordinary" part of the plant. A variety of terms are used to describe the bits that can grow into new plants. For the most part they won't be used here. The important thing is to learn something about the ways in which bryophytes reproduce vegetatively, rather than simply bits of jargon. One widely used expression is vegetative propagule, which is a useful, general term for any piece (large or small) that can produce a new plant vegetatively.
In the form of vegetative reproduction illustrated by Riccia and Sphagnum, the new plants form alongside the original plants. This method produces a fairly slow spread. As noted earlier, some vegetative propagules can be very small and the smaller the fragment the more easily it is carried further away from the parent plant (especially by wind or water). Therefore some of these other means of vegetative reproduction help in more rapid spread of a species and there's more in the DISPERSAL SECTION.
In many cases of re-growth from a vegetative propagule, a new thallose or leafy plant does not simply grow immediately from the propagule. Rather, a protonema first grows out of the propagule and a new thallose or leafy plant develops from the protonema. A protonema is a juvenile, undifferentiated growth form (for example, the first stage in gametophyte development after spore germination). It often resembles a featureless, green wash on its substrate (and you can find out more in the LIFE CYCLE SECTION). In such cases of vegetative reproduction there is a reversion to a juvenile stage before the development of a new thallose or leafy growth![]() .
.
Fragmentation
There are many ways in which part of a bryophyte could get broken off. A wombat starting to dig its burrow scrapes a liverwort colony apart and sends pieces off in various directions. In moist forests you will find curtains of mosses, such as the one pictured here. ![]() Often these will be high up on the forest trees and may be easily broken by storms or animals. That may lead to strands of moss landing elsewhere. In each of the two cases noted here the dispersed pieces will continue growing if their new locations provide suitable conditions. On a more mundane level think of this moss,
Often these will be high up on the forest trees and may be easily broken by storms or animals. That may lead to strands of moss landing elsewhere. In each of the two cases noted here the dispersed pieces will continue growing if their new locations provide suitable conditions. On a more mundane level think of this moss, ![]() growing in a lawn that has been recently mown. You can see the cut ends of the grass blades. The lawn mower may also have cut the moss and flung some fragments further afield.
growing in a lawn that has been recently mown. You can see the cut ends of the grass blades. The lawn mower may also have cut the moss and flung some fragments further afield.
The scenarios in the previous paragraph involve random accidental or deliberate breakages by some other agency, with no real involvement by the bryophytes. It is important to also note that many bryophytes are quite brittle when dry, so making breakage much easier than in the moist state. However many bryophytes (in particular among the mosses and leafy liverworts) naturally develop weakened areas in parts of the gametophyte. Such areas are pre-disposed to breakage and the consequent liberation of particular (rather than random) gametophyte fragments as vegetative propagules.
|

In Campylopus clavatus, a tufted moss, a small apical section of each stem is weakly joined to the rest of the stem. This apical section will break off easily, for example if an animal walks across the moss colony or raindrops hit the colony with sufficient force. You will typically see a colony of Campylopus clavatus with numerous loose apical fragments lying on top of the colony ![]() . Each fragment consists of a very short stem section, with attached leaves, and on the right is a closer view. Such fragments are easily dispersed by wind or water. In fact if you run your finger across such a moss colony you will see those fragments "jump" about, with some landing a little away from the colony. This will happen even if the moss cushion is moist, rather than dry. The moss Pohlia nutans has catkin-like branches which are readily detached from the parent plant. Some hornworts in the genus Megaceros have frilly thallus margins which can fragment readily. In the leafy liverwort genus Pycnolejeunea even a light touch is enough to break off a shoot fragment with several attached leaves. Plants of the Northern Hemisphere leafy liverwort Lejeunea cardotii are prostrate and typically grow as mats on tree trunks or dead wood. Small-leaved branches grow from the large-leaved, main stems of the plant. Those small-leaved branches may be produced in immense numbers, one behind each leaf of a main stem. Furthermore, these small-leaved branches may in turn produce more small-leaved branches. In many colonies of this liverwort the small-leaved branches greatly outnumber the larger-leaved main stems. The small-leaved stems are very fragile and are easily broken off and dispersed as vegetative propagules
. Each fragment consists of a very short stem section, with attached leaves, and on the right is a closer view. Such fragments are easily dispersed by wind or water. In fact if you run your finger across such a moss colony you will see those fragments "jump" about, with some landing a little away from the colony. This will happen even if the moss cushion is moist, rather than dry. The moss Pohlia nutans has catkin-like branches which are readily detached from the parent plant. Some hornworts in the genus Megaceros have frilly thallus margins which can fragment readily. In the leafy liverwort genus Pycnolejeunea even a light touch is enough to break off a shoot fragment with several attached leaves. Plants of the Northern Hemisphere leafy liverwort Lejeunea cardotii are prostrate and typically grow as mats on tree trunks or dead wood. Small-leaved branches grow from the large-leaved, main stems of the plant. Those small-leaved branches may be produced in immense numbers, one behind each leaf of a main stem. Furthermore, these small-leaved branches may in turn produce more small-leaved branches. In many colonies of this liverwort the small-leaved branches greatly outnumber the larger-leaved main stems. The small-leaved stems are very fragile and are easily broken off and dispersed as vegetative propagules![]() .
.
The term deciduous is used to describe such readily-detachable features. Many other leafy liverworts also have deciduous branches and both mosses and leafy liverworts have species with deciduous leaves. In some cases, rather than a whole leaf being deciduous, there's a line of weakness within the leaf so that only a part of the leaf breaks off and disperses readily as a propagule. The leaves in many leafy liverwort species are lobed or with pronounced finger-like projections. In a number of species such lobes or projections break off readily and are dispersed as vegetative propagules.
The northern hemisphere leafy liverwort Gymnocolea inflata has deciduous perianths. A perianth is a protective tube, composed of fused leaves, around an egg-bearing archegonium in a leafy liverwort. Archegonia are part of the SEXUAL REPRODUCTION cycle. In Gymnocolea inflata the perianths around fertilized archegonia do not become deciduous. That's understandable, for otherwise they would not function to protect the developing sporophyte. However, the perianths around un-fertilized archegonia are easily shed. Moreover, the perianths around unfertilized archegonia become more inflated than the ones around fertilized archegonia. So, when shed, each such inflated perianth contains an air bubble which makes the perianth easily carried by water. The perianths of Chonecolea doellingeri, another Northern Hemisphere leafy liverwort, are also very easily dislodged. In this species the deciduous perianths are not of a different form![]() .
.
So far the deciduous fragments have been simply parts of the plant in their expected places. It is also possible for what look like ordinary plant parts to grow in unexpected areas. The moss Pilotrichella flexilis is found in the northern hemisphere. One report from Mexico describes miniature branches growing from the leaves on normal branches and notes that those miniature branches are easily shed. Normally you'd expect branches to grow out from stems or other branches - not from leaf surfaces. The miniature branches (with their own miniature leaves) were essentially of the same structure as normal branches, just much smaller. Any branchlet found growing from a leaf arose from a single leaf cell and a single leaf could give rise to several branchlets. Such branchlets were also found growing from the ordinary branches of the moss. When the branchlets developed on leaves, they did so only from the cells in the basal corner regions, close to where the ordinary leaves joined the ordinary branches. The cells in that area are technically called alar cells and in many mosses are different from other cells in a leaf. There's more about alar cells and their function in the LEAF SECTION. In Pilotrichella the branchlets developed directly from the alar cells, without any intermediate protonemal stage. A number of mosses produce miniature branches as propagules, but the Pilotrichella case was the first time such a development had been observed from moss leaves. The distance between alar cells and the nearby ordinary branch is quite small. But the bryologist who reported this discovery noted that cells which had become differentiated into leaf cells had to de-differentiate and then genes that initiate branch development had to be triggered. The growth of similar miniature shoots (with miniature leaves) has long been known from leaf cells of liverworts in the genus Plagiochila![]() .
.
Gemmae
All the methods of vegetative reproduction described so far have involved the shedding of some part of a well-developed feature. We've seen, for example, that new plants can grow from deciduous branch tips, branches, leaves and miniature stems. Many bryophytes produce gemmae. These are vegetative propagules which are not simply standard parts of the sorts of features just mentioned. Rather they are small, largely undifferentiated growths. Gemmae may be formed as outgrowths from some part of the gametophyte, in which case they are called exogenous gemmae. By contrast, endogenous gemmae are formed within gametophyte cells. Exogenous gemmae are attached to the gametophyte during their development but this attachment weakens as the gemmae mature, allowing the mature gemmae to be broken off fairly easily. Endogenous gemmae are released by the breakdown of the surrounding cell. Exogenous gemmae are more common than endogenous gemmae.
You'll see small, circular cups on the gametophytes of thallose liverworts in the genus Marchantia ![]() . Within these cups you can see some small green discs. Each of those is an exogenous gemma and may get splashed out by a raindrop or washed out by flowing water. The circular cup is called a gemma cup. The great majority of the bryophytes that produce exogenous gemmae don't do so in specialised structures. Instead gemmae are mostly produced simply as outgrowths from some part of the gametophyte – thalli, stems, leaves, rhizoids – depending on species. This photograph
. Within these cups you can see some small green discs. Each of those is an exogenous gemma and may get splashed out by a raindrop or washed out by flowing water. The circular cup is called a gemma cup. The great majority of the bryophytes that produce exogenous gemmae don't do so in specialised structures. Instead gemmae are mostly produced simply as outgrowths from some part of the gametophyte – thalli, stems, leaves, rhizoids – depending on species. This photograph ![]() shows numerous bright green gemmae on the leaves of the liverwort Lethocolea pansa.
shows numerous bright green gemmae on the leaves of the liverwort Lethocolea pansa.
Exogenous gemmae have been found on the rhizoids of over 100 moss species. The fact that rhizoidal gemmae are buried and may function as survival organs (akin to the tubers in various flowering plants) has led to these gemmae also being called tubers. No liverwort or hornwort produces rhizoidal gemmae. Liverworts and hornworts form structures that are also called tubers, but these are not produced on rhizoids. Typically these form as swellings at apices of shoots or lobes. Often the shoot or lobe apex turns towards the earth and grows into the soil, so that the tuberous growth is formed below ground level. Often such a tuberous growth is more a means of survival, rather than a means of reproduction since there is no multiplication of individuals. As with exogenous gemmae, endogenous gemmae may be found in various parts of the gametophyte, depending on the species involved.
Plantlets and a "walking" moss
New plants may grow from the leaf tips of the moss Octoblepharum albidum. In this development rhizoids and, a little later, buds grow at the leaf tips. The buds produce shoots and leaves which, in turn, may produce additional leaf-tip shoots. Such a phenomenon is known in several fern species, where frond tips that come into contact with the ground may root and develop new fern plants. Such ferns are commonly called "walking ferns". The researchers who reported the phenomenon in Octoblepharum noted that in the field such "walking mosses" constituted from 5 to 20 percent of the plants they saw. In contrast, in the mosses they grew in artificial culture over 9 months, the walking variety accounted for about 50% of the plants. In another moss, Leucobryum glaucum, the terminal rosette of leaves may also bear a tuft of rhizoids. New plants may develop on those rhizoids, fall off and then continue growing where they land. Vegetative propagules which look like plant buds grow from cells in at least two species of the thallose liverwort genus Fossombronia, one from Australia and one from South Africa![]() .
.
General comments
|

You've already seen that new plants can grow from a great variety of fragments. In laboratory experiments new plants have been produced from more gametophyte parts, such as archegonial necks and stalks, paraphyses, calyptrae, rhizoid fragments and even the scales on the undersides of thallose liverworts (right) in the family Marchantiaceae. Such results suggest that almost any part of a gametophyte can be induced to produce a new plant.
Thus far the propagules that have been mentioned have been from the gametophyte. In laboratory experiments new plants have been produced from fragments of moss setae. Many mosses have stalked spore capsules and such a stalk is technically called a seta. The seta is part of the sporophyte and is diploid, whereas the gametophyte is haploid. In the haploid stage a bryophyte has one set of chromosomes in each cell nucleus, and in the diploid stage two sets per nucleus. One set of chromosomes comes from each parent and are combined in the process of sexual reproduction. The diploid sporophyte grows from the haploid gametophyte and there's more about the haploid and diploid stages in the LIFE CYCLE SECTION. A setal fragment will give rise to a new gametophyte, but one with two sets of chromosomes in each cell nucleus. Such a gametophyte is therefore genetically distinct from the original gametophyte from which the seta grew![]() .
.
It should not be surprising to find that many bryophytes may reproduce vegetatively in more than one way. For example, there's nothing special about random fragmentation, so there's nothing to stop a gemma-producing bryophyte also shedding such fragments. Bryum argenteum has deciduous shoot tips and also produces rhizoidal gemmae. The moss Octoblepharum albidum, can reproduce vegetatively via gemmae as well as via shoots at leaf tips, as described above. The northern-hemisphere thallose liverwort Blasia pusillus produces two different types of gemmae by quite different mechanisms. This appears to be unique in the liverworts.
Stellate gemmae are produced directly on the thallus surface while ellipsoidal gemmae are produced within long-necked flasks that develop on the thallus surface. Blasia forms a symbiotic association with Nostoc, a CYANOBACTERIAL genus. Each liberated stellate gemma contains Nostoc and, over the summer months, the stellate gemmae germinate to produce fresh, Nostoc-allied thalli. The stellate gemmae do not survive winter. The ellipsoid gemmae do not contain Nostoc and so, when they germinate after liberation, need to form fresh symbiotic associations with free-living Nostoc cells. The ellipsoid gemmae are liberated mainly in late summer to autumn and are able to survive winter. Amongst the mosses there are species which may produce gemmae on the protonema, rhizoids and on the leafy-stemmed plant. The production of one type of gemma neither entails nor precludes the production of another type![]() .
.
This page has been devoted to a description of the various ways in which bryophytes can produce vegetative propagules. That's the first step but for successful reproduction those propagules then need to be dispersed and that's the subject of the DISPERSAL SECTION.
Jargon
This page has used very little technical terminology. As noted earlier, what's important is an understanding of the variety of the processes. However, if you want to look elsewhere for information it will pay to be familiar with some of the terminology. Knowledge of those terms can help you search out relevant information - either from printed or web material.
So, in this section you'll find no additional facts about the nature of vegetative reproduction in bryophytes. There'll be a brief list of various terms used in connection with vegetative reproduction.
Cloning is another way of referring to vegetative reproduction. You may see the expression asexual reproduction used as an equivalent of vegetative reproduction. On the other hand, some people use the term asexual reproduction in a narrower sense - for reproduction via a protonemal stage from some small propagule. That would exclude reproduction via a branching point, as illustrated by Riccia, for example.
Caducous is another way of saying deciduous.
The following terms are used for various forms of vegetative propagules: asexual diaspore, asexual propagule, brood body, bulbil, cladium, surculus, vegetative diaspore. Different writers may have slightly different definitions for a given term.
See also:
Sexual Reproduction
Sexual vs Vegetative Reproduction