![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

Mycorrhizas
The word mycorrhiza is derived from the Classical Greek words for 'mushroom' and 'root'. In a mycorrhizal association the fungal hyphae of an underground mycelium are in contact with plant roots, but without the fungus parasitizing the plant. In fact the association is commonly (but by no means always) mutually beneficial. Through photosynthesis a chlorophyll-containing plant makes simple carbohydrates (using carbon dioxide, water and sunlight). While it is clear that the majority of plants form mycorrhizas, the exact percentage is uncertain, but it is likely to lie somewhere between 80% and 90%. In many of these associations between 10% and 30% of the food produced by the plant moves through to the fungi.
A point about spelling - you’ll often see mycorrhizae as another older spelling of the plural of mycorrhiza.
While the classical Greek word for mushroom is part of the word mycorrhiza,
there are many mycorrhizal fungi which have fruiting bodies other than mushrooms.
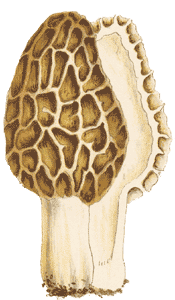
An example is Hydnum repandum ![]() .
It’s superficially mushroom-like (stem and cap), but below the cap there
are spines rather than gills. There’s more about the fruiting bodies of
mycorrhizal fungi later on.
.
It’s superficially mushroom-like (stem and cap), but below the cap there
are spines rather than gills. There’s more about the fruiting bodies of
mycorrhizal fungi later on.
This section gives some basic facts about mycorrhizas. As these associations are important for plant health, they have been extensively researched, both in Australia and overseas. Hence, there is a lot of information available, both on the internet and in printed form. An excellent mycorrhizal website, especially from an Australian perspective, has been created by Mark Brundrett of the University of Western Australia [http://mycorrhizas.info/].It deals mainly with ectomycorrhizas and VA mycorrhizas (both of which will be explained shortly) and gives much detailed information, numerous illustrations, many references and links to numerous other mycorrhizal sites. Further in this section there will be links to specific areas within that site.
Another very useful website, with links to numerous other sites and bibliographies of printed information, is the Mycorrhiza Literature Exchange. [http://mycorrhiza.ag.utk.edu/] The webpage of the new International Mycorrhizal Society (formed in 2002) [http://www.mycorrhizas.org/] may eventually replace the Mycorrhiza Information Exchange.
The printed references given in the button give useful overviews and are the
source of much of the information presented here. I’ll make special mention
of the first of these because it is very recent and with copious references.
![]() It
also has further information on all the topics discussed below and contains
some interesting hypotheses about the evolution of mycorrhizas.
It
also has further information on all the topics discussed below and contains
some interesting hypotheses about the evolution of mycorrhizas.
Different types of mycorrhizas
Physically, there are several forms of mycorrhizas, with different forms of hyphal arrangement or associated microscopic structures. You’ll find micro-photographs of some of these structures in the introductory section of the this site. [http://mycorrhizas.info/#intro]
In vesicular-arbuscular mycorrhizas (or VA mycorrhizas) the fungal hyphae penetrate root cells and form intricately branched, shrub-like arbuscles within the cells and, at times, bladder-like vesicles as well. These associations are often called just arbuscular mycorrhizas, since not all the fungi that are involved form vesicles. There will not be much here about the fungi involved in VA mycorrhizas, not because they are unimportant, but because the fungi that are involved are outside the scope of this website. The VA mycorrhizas certainly are very important and are well-covered here [http://mycorrhizas.info/vam.html]. They are the most abundant type of mycorrhiza and the most ancient. It is likely that these fungi originated between 350 and 450 million years ago and probably played an essential role in the colonization of land by the plants.
The hyphae of ectomycorrhizal fungi form a mantle around the root and
also grow into the spaces between root cells - but do not penetrate the root
cells. The hyphae form a net-like covering, called a Hartig net, around
the cells. You’ll find out a lot more about ectomycorrhizas (and see some very nice photographs of Hartig nets) here [http://mycorrhizas.info/ecm.html]. While the VA mycorrhizas are the most abundant, no more than a few hundred species
of fungi are involved, whereas over 6,000 species of fungi are involved in ectomycorrhizas.
Many of the world’s dominant forest trees form ectomycorrhizas and the
ectomycorrhizal fungi produce many of the commonly seen fungal fruiting bodies
in Australian forests or gardens. To give just two examples, there are numerous
species of Amanita ![]() and Cortinarius
and Cortinarius ![]() in Australia and these two fungal genera form ectomycorrhizal associations with
various native tree species in a wide variety of habitats. There’ll be
more about ectomycorrhizal fungi a little later.
in Australia and these two fungal genera form ectomycorrhizal associations with
various native tree species in a wide variety of habitats. There’ll be
more about ectomycorrhizal fungi a little later.
In ericoid and orchid mycorrhizas [http://www.anos.org.au/groups/newzealand/biology/fungi.htm] the hyphae penetrate the root cells but neither arbuscles nor vesicles are formed.
 Woollsia pungens |
Plants in the order Ericales are the dominant species in many arctic or sub-arctic areas of the northern hemisphere and many of the Ericales are more commonly collectively known as "heaths". The bulk of these plants form ericoid mycorrhizas. Examples of Australian heath plants are the genera Epacris, Leucopogon and Woollsia. These and similar plants are often collectively called the epacrids and ericoid mycorrhizas have been found on many of the Australian epacrid plants.
![]() The
reference given in the button contains much up-to-date information about mycorrhizas
on epacrids and is the source of the various facts about Australian ericoid
mycorrhizas given further on.
The
reference given in the button contains much up-to-date information about mycorrhizas
on epacrids and is the source of the various facts about Australian ericoid
mycorrhizas given further on.
 Thelymitra ixioides a terrestrial orchid |
The terrestrial orchids (of which Australia has many species) form mycorrhizas
and need fungal partners in order for seedlings to survive beyond the germination
stage. ![]() In
at least some species the orchid needs the fungus throughout the orchid’s
life. In these associations the orchid gets the better deal and in effect harvests
the fungus.
In
at least some species the orchid needs the fungus throughout the orchid’s
life. In these associations the orchid gets the better deal and in effect harvests
the fungus.
Other types of mycorrhizas, not yet found in Australia are arbutoid mycorrhizas, ectendomycorrhizas and monotropoid mycorrhizas. In effect, these are variants of ectomycorrhizas.
The ecology and roles of mycorrhizas
In many cases there is an exchange of nutrients between the two organisms, but in some mycorrhizal associations the benefits flow one way rather than both ways, as in the orchid example noted above. Where there is an exchange, the fungus receives some of the carbohydrates photosynthesized by the plant and the plant obtains various inorganic nutrients and trace elements which the mycelium has extracted from the soil.
Photosynthesis provides a chlorophyll-containing plant its source of carbon. Other important nutrients, such as nitrogen, potassium and phosphorus (as well as various trace elements) come from the soil. Usually those other nutrients will be dispersed in the soil, so necessitating the exploration of a large volume of soil if the plant is to get the required amounts of those nutrients. In addition, the non-carbon elements will be present in the soil in various chemical compounds - some of which are readily soluble and easily absorbed by roots whereas others are insoluble (and therefore inaccessible to plant roots). If a plant were to use roots to explore a large volume of soil, it would require a considerable expenditure of energy by the plant. Making use of fungal hyphae saves this expenditure. Moreover, the very fine fungal hyphae can explore much finer cavities than roots and are also capable of exploiting compounds that are inaccessible to roots.
Various mycorrhizal fungi also help protect the associated plants against pathogenic fungi, a number of soil microbes, heavy metals and toxic compounds. Experimental studies have shown that many photosynthesizing plants fail to develop into strong individuals, if deprived of their mycorrhizal partners. All in all, the plants get a fair return on the 10-30% of photosynthesized carbohydrates that pass through to the attached mycelia.
Mycorrhizal associations can be fairly complex, with numerous mycelia (from
more than one fungal species and of more than one type of mycorrhiza) simultaneously
latching onto the roots of one plant. Conversely, a given mycelium may be attached
to more than one plant, perhaps even to plants of different species. This latter
plant-mycelium-plant connection allows the carbon produced by one plant to move
through the attached mycelium to another plant. In this way, a seedling growing
in heavy shade (and therefore unable to make much use of the sunlight) can benefit
from the photosynthesizing activities of some other, better-placed plant. ![]() These
intricate, multiway connections have led some people to refer to the underground
roots-and-mycelia network as the "wood wide web"!
These
intricate, multiway connections have led some people to refer to the underground
roots-and-mycelia network as the "wood wide web"!
In the world there are over 400 species of plants which lack chlorophyll.
These achlorophyllous plants are found in many families and the majority
rely on fungal associates for the carbohydrates which chlorophyllous plants
would produce via photosynthesis. In some cases an entire plant family is achlorophyllous
(such as the Monotropaceae in the northern hemisphere) while in others (such
as the Orchidaceae) only certain genera lack chlorophyll. ![]() You
can find photos and information about achlorophyllous plants here http://staff-www.uni-marburg.de/~b_morpho/imhtopic.html
You
can find photos and information about achlorophyllous plants here http://staff-www.uni-marburg.de/~b_morpho/imhtopic.html
In Australia the orchids Gastrodia sesamoides (the Potato Orchid), Dipodium punctatum (the Spotted Hyacinth Orchid) and the two species of the genus Rhizanthella (Rhizanthella gardneri and Rhizanthella slateri) lack chlorophyll. The Rhizanthella species are unusual because they are underground orchids, whereas both Gastrodia and Dipodium produce above-ground flower spikes. The fungi involved in these associations are quite varied, both in terms of species and lifestyles. For example the species of Gastrodia (both in Australia and overseas) are associated with species of Armillaria (a genus of virulently parasitic fungi, which the orchids manage to keep under control). The Western Australian underground orchid Rhizanthella gardneri forms an orchid mycorrhiza with a fungus that also forms ectomycorrhizas with Melaleuca (one of the Australian native Tea Trees). Incidentally, this also shows that some fungi can form more than one type of mycorrhiza. The eastern Australian species, Rhizanthella slateri, pictured here, has been rarely seen. It was discovered in 1931 on Alum Mountain, in Bulahdelah on the New South Wales north coast, and these photos (taken in 2002) are also of specimens growing on Alum Mountain. The fungal associate of this species is unknown.
 Rhizanthella slateri |
 Rhizanthella slateri |
All such orchids are directly parasitic on the associated fungi. The carbohydrates that end up in Gastrodia and at least the Western Australian Rhizanthella originated in other living plants, so you could also look at these orchids as being indirectly parasitic on other plants.
 Orobanche australiana |
Not all the achlorophyllous plants have fungal associates. Some attach directly
to the roots of host plants and the best known are probably the Broomrapes,
which constitute the genus Orobanche. Here is an Australian example,
Orobanche australiana, and this link http://www.parasiticplants.siu.edu/Scrophulariaceae/Orobanche.Gallery.html leads to a photo gallery of Orobanche species from around the world.
Another example is the genus Balanophora ![]()
![]() .
Both Balanophora and Orobanche are flowering plants, but are often
mistaken for a fungi.
.
Both Balanophora and Orobanche are flowering plants, but are often
mistaken for a fungi.
The mycorrhizal associations formed by a particular plant need not be static. Over time, the fungi that are involved can change. Some may associate only with young plants, others come into the picture only when the plant is a few years old - while some may stick around and it is possible that, depending on conditions, there are some which come and go. A study of the ericoid mycorrhizas on several epacrid species in the south-west of Western Australia showed that the mycorrhizas were seasonal. The area has a Mediterranean climate with cool, wet winters but hot, dry summers. The mycorrhizas are formed on the very fine hair roots that grow out from the main roots. In summer many of the hair roots die but regrow, and become mycorrhizal, in the wet winter months. Similar studies around Sydney, not subject to a Mediterranean climate, didn’t show this die-regrow pattern and studies in the northern hemisphere also show year-round persistence of mycorrhizal roots in most species. In summary it seems that, except in very dry conditions, ericoid mycorrhizas are present and functioning all year round.
Finally, the formation of the mycelium-root connection is not just a simple
matter of fungus-meets-plant, fungus-hooks-onto-plant. ![]() There
are soil bacteria, known as mycorrhizal helper bacteria, which promote
the formation of mycorrhizas.
There
are soil bacteria, known as mycorrhizal helper bacteria, which promote
the formation of mycorrhizas.
You’ll find out much more about the roles of mycorrhizas here [http://mycorrhizas.info/roles.html].
Which plants form mycorrhizas?
![]() It
has been estimated that at least 80% and perhaps up to 90% of the world's plants
form mycorrhizas of one form or another and you will find mycorrhizal associations
from well-watered forests to the arid areas.
It
has been estimated that at least 80% and perhaps up to 90% of the world's plants
form mycorrhizas of one form or another and you will find mycorrhizal associations
from well-watered forests to the arid areas.
The table on this page [http://mycorrhizas.info/ozplants.html] gives the mycorrhizal status of numerous genera of Australian plants. Many of the imported garden trees in Australia are also mycorrhizal, for example beeches, oaks, firs and pines.
 Banksia praemorsa |
Though a great many Australian plants form mycorrhizas, especially VA mycorrhizas, there are some noteworthy exceptions. For example, many genera in the Proteaceae (which includes the widespread genera Banksia, Grevillea and Hakea) do not form mycorrhizas. Even in artificial conditions, when attempts are made to force mycorrhizas these genera seem to actively resist mycorrhizal formation.
While 80-90% of plants form mycorrhizas, the proportion of mycorrhizal plants in a particular habitat may be markedly different from the percentage range just given. Depending on the plant composition of an area the proportion of mycorrhizal plants may be virtually 100% or less than 50%. Given what was just said about the Proteaceae, just think of an area of Australian bushland dominated by Grevillea and Hakea with just a few eucalypts and a scattering of other plants.
Liverworts are another group of plants with fungal associates. Though liverworts are plants they lack both roots and the internal nutrient conducting systems found in the ferns and flowering plants. Liverworts have anchoring filaments, called rhizoids, which are superficially root-like in appearance but, unlike roots, do not draw water or minerals from the soil. Instead, water and minerals are absorbed through the liverwort surfaces. So, strictly speaking, liverworts cannot form mycorrhizas, since these have been defined as requiring roots. However, a number of liverworts form fungal associations that physically resemble mycorrhizas, with fungal hyphae found in the liverwort rhizoids. The functions of some of these liverwort-fungus associations still need to be determined.
In the northern hemisphere heath plants (in the family Ericaceae) and some groups of liverworts can form associations with the same group of fungi. Fungi that are either the same as (or very similar to) the northern hemisphere ones have also been observed in ericoid mycorrhizas with Australian epacrids as well as in mycorrhiza-like associations with some Australian liverworts. One suggestion is that the liverwort rhizoids serve as a source from which the fungi can colonize plant roots. However, the nature of the association (if any) between liverworts, fungi and northern hemisphere heaths (or Australian epacrids) is unknown but is being investigated.
Almost all liverworts contain chlorophyll and so produce simple sugars through
photosynthesis, just as the majority of other plants do. The genus Cryptothallus,
found in the northern hemisphere, is an exception and lacks chlorophyll. ![]() As
in the case of the achlorophyllous orchids mentioned above, Cryptothallus
relies on its fungal partner for nutrients.
As
in the case of the achlorophyllous orchids mentioned above, Cryptothallus
relies on its fungal partner for nutrients.
Which fungi form mycorrhizas?
The fungi that form mycorrhizas are quite varied. Ectomycorrhizal fungi produce
many of the easily seen fruiting bodies that you commonly come across. There
are many mushroom-producing ectomycorrhizal fungi and examples of Amanita
and Cortinarius were given earlier. These are large mycorrhizal genera,
with many species worldwide. While there are many native Amanita species
in Australia, there are also some introduced species. For example, Amanita
muscaria ![]() (commonly called the Fly Agaric) and Amanita phalloides
(commonly called the Fly Agaric) and Amanita phalloides ![]() (known as the Deathcap [see DEATHCAP SECTION])
have been introduced unintentionally. You’ll commonly see the first of
these near pine or birch trees and the latter near oak trees. However, in Tasmania
Amanita muscaria has been found growing in native Nothofagus forests
- with not a pine or birch tree anywhere to be seen. Amanita muscaria
has also been introduced to New Zealand, where it has been found in native forests
with Kunzea and Leptospermum. While some fungi may have preferences
for particular plants, the Tasmanian and New Zealand reports show that at least
some mycorrhizal fungi can take up new partners.
(known as the Deathcap [see DEATHCAP SECTION])
have been introduced unintentionally. You’ll commonly see the first of
these near pine or birch trees and the latter near oak trees. However, in Tasmania
Amanita muscaria has been found growing in native Nothofagus forests
- with not a pine or birch tree anywhere to be seen. Amanita muscaria
has also been introduced to New Zealand, where it has been found in native forests
with Kunzea and Leptospermum. While some fungi may have preferences
for particular plants, the Tasmanian and New Zealand reports show that at least
some mycorrhizal fungi can take up new partners. ![]() There
has also been a report of Amanita phalloides associating with eucalypts
in Canberra (but this requires more investigation) and there are records of
that species in a eucalypt plantation in Tanzania and near eucalypts in Algeria.
There
has also been a report of Amanita phalloides associating with eucalypts
in Canberra (but this requires more investigation) and there are records of
that species in a eucalypt plantation in Tanzania and near eucalypts in Algeria.
Apart from Amanita and Cortinarius there are many more species
of mushroom-producing ectomycorrhizal fungi in various genera - Hebeloma
![]() ,
Inocybe, Lactarius
,
Inocybe, Lactarius ![]() ,
Paxillus, and Russula to give a few examples of commonly seen
genera.
,
Paxillus, and Russula to give a few examples of commonly seen
genera.
As noted at the beginning of this mycorrhizal section, there are numerous non-mushroom,
ectomycorrhizal fungi and you’ve already seen the example of Hydnum
repandum, which was pictured right at the start. The majority of truffle-like
fungi are mycorrhizal and examples of these are the introduced species Melanogaster
ambiguus ![]() (which is found growing near introduced oak trees), this native species of Setchelliogaster
(which is found growing near introduced oak trees), this native species of Setchelliogaster
![]() and Peziza whitei
and Peziza whitei ![]() ,
another endemic Australian species. Some of the coral fungi, many boletes, such
as the native Phlebopus marginatus
,
another endemic Australian species. Some of the coral fungi, many boletes, such
as the native Phlebopus marginatus ![]() and the introduced Suillus luteus
and the introduced Suillus luteus ![]() (which is found under non-native pine trees) and a few powdery, superficially
puffball-like fungi (such as Pisolithus
(which is found under non-native pine trees) and a few powdery, superficially
puffball-like fungi (such as Pisolithus ![]() and Scleroderma
and Scleroderma ![]() )
are mycorrhizal.
)
are mycorrhizal.
The earthstar-like Astraeus hygrometricum ![]() (introduced to Australia) is known to be ectomycorrhizal with various northern
hemisphere tree species.
(introduced to Australia) is known to be ectomycorrhizal with various northern
hemisphere tree species.
Species of Morchella ![]() can grow quite well as saprotrophs, but some overseas studies
suggest that the species of Morchella may sometimes form ectomycorrhizas.
can grow quite well as saprotrophs, but some overseas studies
suggest that the species of Morchella may sometimes form ectomycorrhizas.
The corticioid fungi commonly produce their flat, sheet-like fruiting bodies on the underside of dead wood, twigs and similar plant litter on the forest floor. It would be excusable to suppose that these are all saprotrophic fungi, feeding on the dead plant matter on which the fruiting bodies are formed. That is often the case but in the northern hemisphere there are about a hundred species of corticioid fungi that are mycorrhizal, many of them in the genus Tomentella. A probable mycorrhizal Tomentella has been found in Australia recently, but the majority of Australian corticioid fungi are poorly studied.
The mycorrhizal nature of a few of those hundred corticioid species in the
northern hemisphere has been known for a long time, but in most cases the mycorrhizal
evidence was discovered in the late 1900s. These corticioid genera contain confirmed
ectomycorrhizal species: Amphinema, Byssocorticium, Byssoporia, Piloderma,
Pseudotomentella, Tomentella, Tylospora. Tylopsora fibrillosa is
not known from Australia and the photo ![]() was taken in a Sphagnum bog in Estonia.
was taken in a Sphagnum bog in Estonia. ![]() The
whitish bloom growing over the Sphagnum plants is the Tylospora
fruiting body and is lined with the spore-producing basidia. These corticioid
genera are suspected to contain ectomycorrhizal species: Athelia, Lindtneria,
Tomentellopsis, Trechispora.
The
whitish bloom growing over the Sphagnum plants is the Tylospora
fruiting body and is lined with the spore-producing basidia. These corticioid
genera are suspected to contain ectomycorrhizal species: Athelia, Lindtneria,
Tomentellopsis, Trechispora.
There are more photographs of fruiting bodies of ectomycorrhizal fungi here [http://mycorrhizas.info/ecmf.html].
Many of the VA mycorrhizal fungi do not produce fruiting bodies, the spores often being produced on hyphae in the soil near plant roots. Where fruiting bodies are formed they are usually more-or-less spherical in shape and truffle-like, in that they are hidden in the soil or leaf litter. Many fruiting bodies are small, only a millimetre or so in diameter, though some can get to a couple of centimetres or more in diameter. The fungi are neither ascomycetes nor basidiomycetes and are outside the scope of this website.
Identification of the fungi present in ericoid mycorrhizas is mostly based on the physical, chemical or DNA analyses of the hyphae found in roots, since fruiting bodies are rarely found. Understandably, there is still very much to be learnt about the fungi involved. The best known ericoid mycorrhizal fungus, Hymenoscyphus ericae, is an ascomycete that produces small disc-like fruiting bodies up to 1mm in diameter. However, it appears the fruiting bodies have never been seen in the wild and have been produced in laboratory experiments, only with considerable difficulty. While there is still much ignorance about the species of fungi involved, the various chemical, physical and DNA studies indicate that there is considerable similarity between the northern and southern hemisphere fungi that form ericoid mycorrhizas.
Orchids form mycorrhizas with various macro and micro fungi. The association with the mushroom genus Armillaria was mentioned above, but a number of orchids are known to form associations with macrofungi in the genera Ceratobasidium, Sebacina, Thanatephorus, Thelephora, Tomentella and Tulasnella. Many of these form inconspicuous fruiting bodies, sometimes being no more than powdery or cobwebby coatings on forest litter.
A thousand words
Since a picture is worth a thousand words, here’s a simple diagram (plus a few words) which portrays some of the complexities described earlier.
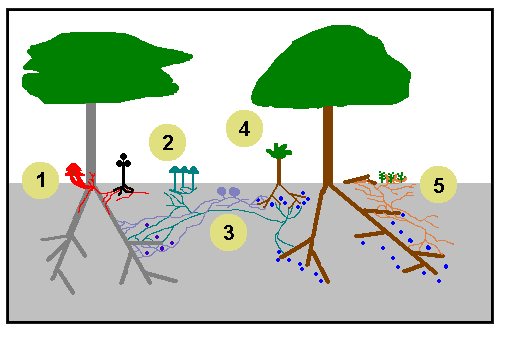
So, what’s going on....
 |
Well at 1 we have an achlorophyllous plant (in black) using a parasitic fungus (in red) as its source of carbon. Notice that the red hyphae have penetrated a tree and some red mushrooms have been formed at the base of the trunk. |
 |
At 2 there are some mushrooms that have arisen from a mycorrhizal fungus. Note how the associated mycelium is linking several plants. |
 |
You can see the buried fruiting bodies of a mycorrhizal, truffle-like fungus at 3. In this case the mycelium connects just two plants. |
 |
The young plant at 4 is growing in the shade of a mature plant, but it is connected to two mature plants by a couple of mycorrhizal mycelia. Notice the numerous dark blue Mycorrhizal Helper Bacteria. |
 |
The mycelium of yet another mycorrhizal fungus has formed corticioid fruiting bodies on fallen twigs and mosses at 5. |