![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

Mistletoes - evolution
Mistletoes of Australia and the World: Evolutionary Origin
The fundamental question about evolution of mistletoes is, of course, how did they come to be aerial parasites on tree branches? However there are other interesting aspects of mistletoe evolution, including changes in floral structure relating to pollination, and to the structure of the fruit.
Origin of the mistletoe habit
How did mistletoes become aerial parasites? Two scenarios seem possible. Firstly, they could have climbed trees first, then become parasites. In other words, mistletoes could have evolved from free-living ancestors which were epiphytic on tree branches in humid forests. Alternatively, they could have become parasites first, then climbed trees. In other words, mistletoes could have evolved from terrestrial root-parasites by somehow adapting as parasites on tree branches.
|
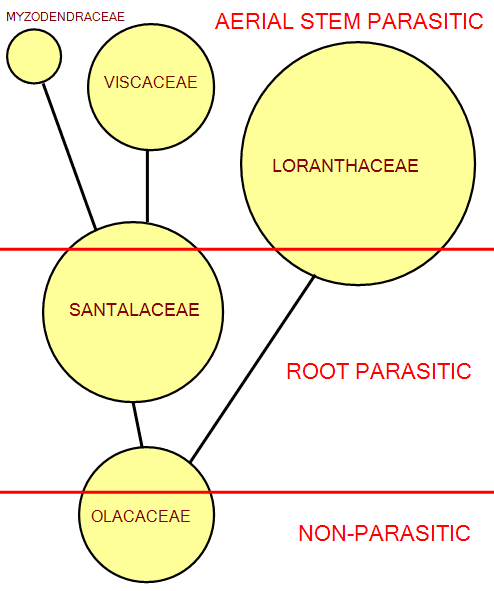
All of the available evidence supports the second hypothesis. Among the order Santalales, to which the different groups of mistletoes belong, the least specialized, presumably ancestral members are free-living trees and shrubs. However the order also includes derived genera which are terrestrial root-parasitic trees and shrubs, and it appears that adoption of root-parasitism has been a central feature in evolution in the Santalales. Finally, the aerial parasites (mistletoes) are found in four distinct families, and represent the most advanced state in the order. It appears that the mistletoe habit has arisen at least four times in the order, from root-parasitic ancestors. Thus the basal family Olacaceae has mostly free-living members, along with a lesser number of root-parasites, whilst the Santalaceae has mostly root-parasitic members, along with a few aerial parasites (diagram left). In the Loranthaceae we find three primitive monotypic genera of root-parasites, all the other species being aerial parasites (see box). All members of the advanced families Viscaceae and Myzodendraceae exhibit the mistletoe habit.
|
The origin of the aerial habit from root parasitism has been explained in two ways. Firstly, it could have resulted from a vine-like intermediate which initially grew in the ground, but formed haustorial connections to tree stems, and eventually lost its contact with the ground. When the root zone of the embryo itself became modified into a haustorium, the plant would be an aerial parasite.
The second possibility is that there was a more direct transfer from the soil to the tree branch. This would depend on a fruit/seed structure suitable for germination on the branch. The plant’s roots may have depended to some extent on accumulated detritus on the tree branches, but presumably also formed haustorial connections to the host’s stems. Again, when the root zone of the embryo itself became modified into a haustorium, the transition to aerial parasitism would be complete.
In both of these scenarios, it follows that the ancestral haustorial system in the aerial parasites would be based on epicortical runners. The more complex primary haustoria, lacking runners and secondary haustoria, would be the final derived state. Indeed, it does appear that epicortical runners are more common among the less specialized mistletoe genera, although they seem to have been widely retained in rainforest mistletoes, and often lost in mistletoes of open and arid habitats.
There is some evidence which favours the second of these scenarios. There are no transitional vine-like states among the Santalales. On the other hand, in the Loranthaceae some genera such as the Australian Muellerina ![]() have seedling stages in which the first epicortical runner is rather inflexibly fixed in position relative to the seeding stem, and this seems to be a transitional step to a condition in which epicortical runners can develop freely regardless of the position of the seed on the branch, and thus favour more efficient establishment.
have seedling stages in which the first epicortical runner is rather inflexibly fixed in position relative to the seeding stem, and this seems to be a transitional step to a condition in which epicortical runners can develop freely regardless of the position of the seed on the branch, and thus favour more efficient establishment.
|
It seems, therefore, that mistletoes evolved from terrestrial root-parasitic progenitors through a sequence in which bird-dispersed seeds were lodged and grew on tree branches (presumably in humid forests), with their roots functioning as epicortical runners and able to make haustorial connections to the tree’s stems. Subsequent adaptations would have involved changes in fruit structure (making bird dispersal more dependable), embryo structure (producing a primary embryonic haustorium, and favouring spontaneous germination), and finally specialization of the haustoria themselves, depending on the demands of the habitat.
Some studies suggest that aerial parasitism has originated more than once in some families within Santalales. For example, it is thought that the mistletoe habit may have originated three times in the Santalaceae (including in the progenitors of Viscaceae). In the Loranthaceae different patterns of embryo structure, germination (see box) and establishment suggest four separate events leading to aerial parasitism (1) in the 12-chromosome, predominantly tropical mistletoes of the Asian-Australasian region of the Old World, (2) in the 11- and 9-chromosome mistletoes of the Old World, (3) in the 8-chromosome mistletoes of the New World, and finally (4) in the relictual Notanthera heterophylla of temperate South America. So, including the event leading to Myzodendraceae, the mistletoe habit may have originated independently at least eight times in the Santalales.