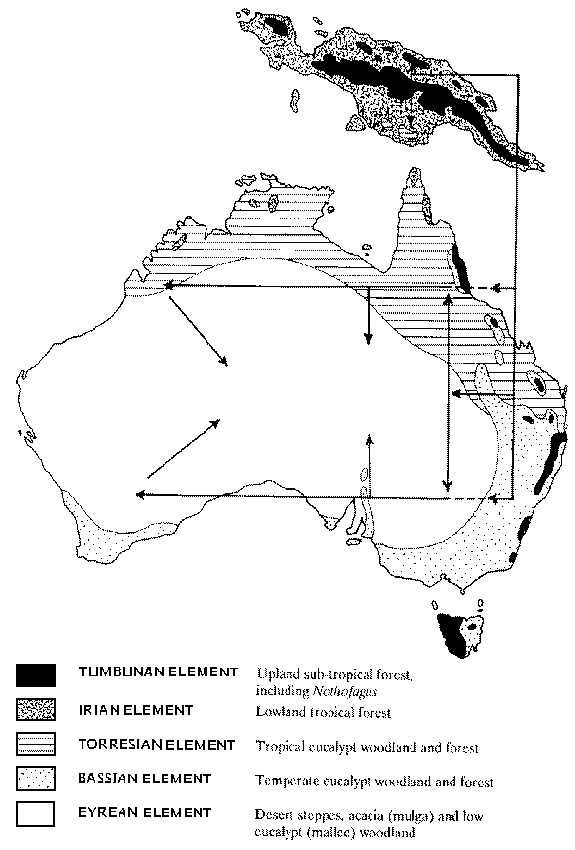
Nancy T. Burbidge Memorial Lecture, 1989Origins, Radiations and Sifting in the Australasian Biota - Changing Concepts from New Data and OldRichard Schodde (From ASBS Newsletter Number 60, September 1989) The Nancy Burbidge Memorial Lecture is held regularly in conjunction with the Societys Annual General Meeting to commemorate this outstanding Australian botanist of the post-war era. Our invited speaker for 1989 was Richard Schodde, who currently curates the wildlife collection at the CSIRO Division of Wildlife and Ecology in Canberra. Dick's connection with plants has been far closer than his present position may indicate. He started his career as a botanist in Adelaide, where he revised Millotia (Asteraccae) for his Honours Science degree under Hansjoerg Eichler. From there, he joined the CSIRO Division of Land Research for whom he made extensive collections of plants in Papua-New Guinea. He revised Pittosporaccae in Papuasia, then returned to Adelaide to complete a Ph.D. on the systematics of the Atherospermataceae. After returning to CSIRO in Canberra, he eventually found his present post with the Wildlife Division, where he made a profession of his long-standing interest in birds. Dick has become an authority on bird systematics and has written and edited several monographic books including the "Readers Digest Complete Book of Australian Birds". It is a very great pleasure for me to be here today to give the Nancy Burbidge Memorial Lecture, for three different reasons. First, and foremost, it allows me to honour my friend and colleague, Nancy Burbidge, and pay homage to her tireless productivity. We were not only fellow travellers in plant systematics and biogeography in the 10 years that I worked at the National Herbarium in Canberra, but also active members in the local National Parks Association after hours - at least Nancy was active. Secondly, it gives me a chance to remeet and mix it with old friends that I have seen little of since I moved from systematic botany to ornithology and the curatorship of the Australian National Wildlife Collection 18 years ago. And thirdly, it allows me to share with you the zoological side of the dramatic changes that have taken place in our understanding of the origins and evolutionary radiation of Australia's biota over the last 20-30 years. To appreciate the extent of the changes, it may help to recapitulate the earlier views of biotic origins in Australia and the reasons for them. For the flora, Nancy Burbidge's own great summary in 1960 of the phytogeography of the Australian region was the apotheosis of aconventional interpretation that had continued from the time of J.D. Hooker. This interpretation, as I'm sure you are all aware, saw the Australian flora as comprising three principal elements: first, an old autochthonous element of scleromorphic proteads, myrtles, epacrids, and many other families that make Australia's landscape so different from the rest of the world; secondly, an Antarctic element of cool temperate myrtle beech-podocarp rainforest and alpine vegetation in extreme south-east Australia and Tasmania; and thirdly, an Indo-Malayan element, comprising the tropical and subtropical rainforests of the north and east coasts. Both Antarctic and Indo-Malayan elements were perceived as recent immigrants, the Antarctic from somewhere in the south and the Indo-Malayan from the Eurasian tropics via New Guinea. In this broad context, the New Guinean flora was almost entirely Indo-Malayan, according to the analyses of van Steenis; even its alpine flora was derived from east Asian sources. New Zealand's, by contrast, was prevailingly Antarctic. What of the fauna? In this lecture I shall confine myself to the land vertebrates - for two reasons. First, I am far more familiar with them. And secondly, most invertebrate phyla and classes are still so poorly known taxonomically that biogeographic conclusions of any significance are difficult to reach. The vertebrate faunas of Australia, New Guinea and New Zealand all include frogs, reptiles, birds and mammals, although New Zealand's complement is far more sketchy than those of the other two. Sharing some 370 species roughly equally between them, Australia's and New Guinea's amphibian faunas are distinctive rather than diverse by world standards. They have no salamanders or sirens, only frogs, and of them they share only 4 of the world's 23 or so families. One family comprises the austral frogs, Myobatrachidae, which are endemic to the region, dominate in Australia, just reach southern New Guinea and have as their nearest relatives the leptodactylid frogs of South America. Another comprises the hylid tree frogs in the form of an endemic regional off-shoot of this nearly cosmopolitan family; its species are the second most abundant in both Australia and New Guinea. The third family, Microhylidae, is particularly diverse in lower and mid-montane New Guinea, and has outliers in the Atherton bloc rainforests of north-east Queensland; elsewhere it is generally pan-tropic. The last family, Ranidae, comprises the true frogs and is cosmopolitan, with greatest diversity in Eurasia. Its 21 species in New Guinea and one in north Australia are members of two genera that are widespread in Eurasia and the Pacific. New Zealand has only three species of frogs, all members of a primitive endemic family, Leiopelmatidae, which is related not to any other Australasian family but to relictual ascaphids in north-west America. Why, you might wonder, am I going into such detail about the distributions of so insignificant a group of animals. The reason is simple. Amphibians are intolerant of salt water, and cannot fly over physical barriers or travel far; among vertebrates they are one of the best indicators of past geographical connections among terrestrial biota. Fresh-water turtles and tortoises are too. They are missing from New Zealand and only a handful survive in Australia and New Guinea. Those that do reveal a pattern repeated again and again in land reptiles, birds and mammals. Three families are involved. One, the pitted-shell turtles, is represented by an endemic species that is the families' sole survivor from Mesozoic times. Another, widespread in Afro-Asia, is represented by a single Malesian species that just reaches New Guinea. The third, comprising 20 species shared between Australia and New Guinea, occurs elsewhere only in South America and belongs to a suborder of side-necked tortoises found today only on the southern continents and Madagascar. Land reptiles are far more diverse in Australia and New Guinea, both lands sharing about 910 species; Australia is decidely the richer with some 680, reflecting the opportunities for evolutionary radiation in its warm and zone. Five of the world's 16 families of lizards are represented on both lands, plus five of the 11 families of snakes; legless lizards, geckos, dragons, goannas, skinks, blind snakes, pythons, file snakes, and solid-toothed and frontfanged snakes. Only one family, the small one of the legless lizards, is endemic; and the closest extra-continental relatives of it and the other families all range through the Malesian archipelago and Afro-Asia. What is not so obvious is a pattern of bicentric distribution between the families. Either they are centred in Afro-Asia and reach Australo-Papua on a comparatively small scale. Such are the gekkonine geckos, skinks, pythons, blind snakes and solid-toothed snakes, which just trickle in. Or, like legless lizards, diplodactyline geckos, dragons, goannas and highly venomous frontfanged snakes, they are centred in Australo-Papua and radiate through Afro-Asia in a more limited way. Coincidentally, the closest family relatives of the dragons and goannas are the American iguanas and Gila Monsters, respectively. New Zealand's reptile fauna is as extraordinary as it is almost non-existent, comprising just 11 primitive endemic diplodactyline geckos, 18 skinks derived from Australian stocks, and the Tuatara, the world's single living rynchocephalian dinosaur. With about 980 land and freshwater species, the Australian and New Guinean bird fauna is very rich and third only to those in South America and Africa. I have grouped the Australo-Papuan bird faunas together because their dominant families - parrots and cockatoos, frogmouths, wood kingfishers, honeyeaters, acanthizid warblers, and thick-head flycatchers are shared by and centred in the region. At the present imperfect state of knowledge, no major New Guinean families are missing from Australia; even birds-of-paradise are here. And the only three Australian families missing from New Guinea - the plains wanderers, lyrebirds and primitive scrub-birds - are found nowhere else either. Outside the region, the relationships of the many orders and families of Australo-Papuan birds lie numerically with Eurasia. Among non-passerines - that is, all the orders of birds that do not sing - most of the Australo-Papuan wading birds, ducks, fovas, birds-of-prey, rails, pigeons, and nocturnal birds are outlying members of groups more diverse in Eurasia. Most passerines or songbirds, moreover, such as the Australo-Papuan robins, flycatchers, wrens, warblers, thrushes and babblers, resemble, in their morphology, the robins, warblers and flycatchers etc. of Eurasia. The one further point that I would like to draw to your attention here, once more, is the sniff of a bicentric pattern of distribution in the non-passerine orders. Again and again it crops up, in the hawks, Magpiegeese, mould-builing hens, stiff-tailed ducks, plains wanderers, and parrots. Genera in these orders that are endemic to Australo-Papua are either obscure in their intercontinental relationships or have links with vicariant groups in South America. What of the New Zealand land and freshwater bird fauna? Birds are that island's most diverse group of vertebrates, yet, with only about 80 species, they are a depauperate and patchy mix of a few very old endemics with no close relatives - the kiwis and moas, nestorine parrots and rifleman wrens - plus equally few endemic genera and families derived from Australo-Papuan stocks - the kakariki parrots, mohouine warblers and callaeatid crowshrikes. Added to them is a considerably greater number of contemporary Australian species that have evidently been blown across the Tasman Sea to colonise the islands. The New Zealand mammal fauna is so depauperate that it can be virtually discounted: no egg-laying monotremes, no marsupials, and no placental mammals are present other than two insectivorous bats. One of the bats is closely related to species in Australia and New Caledonia, and the other, curiously, represents an endemic family related to the cosmopolitan mastiff bats. The Australian and New Guinean mammal faunas, although considerably richer with some 350 species between them, are still poor and patchy by world standards. Yet they include the egg-laying monotremes and groups of marsupials found nowhere else. Both monotremes (two echidnas) and marsupials occur in New Guinea as well as Australia, although not with such diversity. The only two groups of placental mammals in the region rodents and bats - are also shared by both land masses and are moderately rich in species and genera. Here again is that division in distribution patterns, one set directed towards Eurasia, the other to South America. Both groups of placental mammals in Australo-Papua are but two of the many groups of placentals found throughout the rest of the world; and both, moreover, are members of families that are more diverse in Eurasia. The monotremes, however, have no extant relatives anywhere while the marsupals, rich in diversity and endemism, are represented outside only by the didelphid opossums in South America. How did vertebrate zoogeographers up to the 1960's interpret these patterns? Led by Bernard Rensch, George Gaylord Simpson, Ernst Mayr and P.J. Darlington, they first grouped the Australian and New Guinean faunas together as one major element among the world's faunas. Their common yet distinctive mix of frogs, sidenecked turtles, snakes, birds, marsupials and rodents compelled this - and rightly so. New Zealand was virtually discarded, an island waif with faunal flotsam and jetsam that had washed up on it over time and had evolved divergently in its own way. Secondly, and this is the important point, the zoogeographers played the numbers game. Reptiles and birds were the dominant groups in the fauna, and their members had prevailing links with Eurasia. Bats and murid rodents, which together roughly equal the marsupials in numbers of species, were also Eurasian; and so too the ranid frogs. If they could reach antipodean Australia-New Guinea from Asia, as they probably did, so could the rest. Out of this developed a theory of dispersal to explain the origin of Australasian vertebrates. It rested on two particular circumstances and one principle. The two circumstances were, first, the patterns of distribution that I have described; and, secondly, the steady state of the earth's crust, its continents and archipelagos in supposedly fixed position. Over Tertiary time, the Indonesian archipelago had served as the stepping stones for one wave of animals after another dispersing into Australia from the Afro-Asian cradle of vertebrate evolution. Those that came first diverged farthest - the austral frogs, side-necked turtles, emus and parrots, and monotremes and marsupials. And those that came last, presumably within the last million or two years, changed least. Among them were (and are) the ranid frogs, solid-toothed snakes, single species of Eurasian larks, pipits, thrushes and sunbirds, and the true rats, and many insectivorous bats. The principle that contributed to this theory was one of competitive succession. According to it, more primitive less competitive groups were constantly being pushed out and replaced by more advanced and successful groups in the Afro-Asian evolutionary cradle. This was used to dismiss the one great distributional anomaly: the uniquely shared occurrence of the sidenecked turtles, marsupials and flightless ratite birds on the tips of the southern continents. Competition had consigned them to refuges in the antipodes. Similarities in form between the ratites, marsupials and others on the different southern continents, moreover, were often explained also not by common ancestry but by evolutionary convergence through occupation of equivalent niches. As far as Australia was concerned, immigration was by two pathways. One, via the Lesser Sundas to the north-west, was older and led to the development of endemic groups such as the austral frogs, diplodactyline geckos, honeyeaters and marsupials in the old autochthonous Australian flora, perhaps coevally. The other, via New Guinea to the north, was newer, perhaps Plio-Pleistocene in age, and had led to the ingress of rainforest taxa down the east coast, along with the Indo-Malayan elements of the flora. The scenarios pieced together for the origin of Australia's phanerogam flora and vertebrate fauna matched closely and gained from mutual support. Within Australia, late Tertiary climates zoned the biota concentrically, creating a tropical or Torresian province across the north, a temperate or Bassian province around the south, and an Eremaean or Eyrean province in the centre. Again the phytogeographers and vertebrate zoogeographers agreed. Both old autochthonous and lndo-Malayan rainforest elements were included, undistinguished, in the tropical Torresian province. What a comfortable theory it was, fitting the systematic and biogeographic ideas of the day like a hand in a glove. The only biogeographer to challenge it seriously was the Queensland botanist, D.A. Herbert. With rare insight he claimed that Australia's rainforests were not new but old and that elements of the autochthonous flora had evolved out of them by processes of ecological sifting. Leo Croizat also challenged the theory, as he challenged everything, in his world view - but I don't want to get sidetracked into his Genorheitron. Vertebrate zoogeographers, however, found little to question and were content to leave the running to dispersalists like Mayr, Darlington and their disciple Allen Keast. Cladism and its biogeographic sister, vicariance, had not yet taken root; and the implications of vicariance in the global distribution of marsupials, parrots, side-necked tortoises and austral frogs went unremarked. Australia was still seen as something of a biotic vacuum, a bottomless cup waiting to be filled with everything that Eurasia could pour into it. The idea that Australia might have also had a vertebrate fauna, as well as a flora, as old as any in the world and had contributed colonists from its shores to Eurasia in reverse was barely considered. To be sure, Mayr had pointed out that an Australian cuckoo-shrike here and a whistler there had established itself in South-East Asia; but to think that whole groups of Eurasian vertebrates, such as its front-fanged snakes and song bird families, could have stemmed from Australian ancestors was thinking the unthinkable. How much it was all to change and how soon. Literally overnight, continental drift took the ground from beneath it. The shifting position of the earth's continental plates through time, by whatever process, turned from fiction to fact in the 1960's and early 1970's. When Australia was supposed to be getting its first stocks of Eurasian vertebrates and perhaps angiosperms in later Cretaceous to early Tertiary times, some 120-60 million years ago, it was nowhere near its present position but several thousands of kilometres south beside Antarctica. It was, as is now well known, a fragment of the vast southern supercontinent Gondwana which was positioned just north of the south pole in late Palaeozoic and Mesozoic times. Separating Gondwana from the ancestral Eurasian continent, Laurasia, was the Tethys Sea, a barrier that seems to have been much wider and more effective than the island-dotted Indonesian seas today. Gondwana began breaking up about 120 million years ago, beginning with the rifting of Africa and South America and the splitting off of India. New Zealand broke away next from east Gondwana 80-85 million years ago, followed by South America from west Gondwana early in the Tertiary at 60-70 million years. Last to split were the final remnants, Australia and Antarctica, some 50-60 million years ago. Australia remained close to Antarctica for several million years before finally commencing its northward drift to its present position about 40-45 million years ago, into the Oligocene. New Guinea began to form then on, along the northern edge of the Australian continental plate, developing in two parts. One part was the northern rim of the Australian plate itself and the other a string of islands off the north-east coast, away from Laurasia. The islands and mainland only came together towards the end of the Tertiary, throwing up the mighty central cordilleras there in Plio-Pleistocene times and giving New Guinea its present form. This knowledge affected the Eurasian immigrant theory for vertebrates in two ways. First, it drew attention to a possible Gondwanan origin for such primitive - and so presumably ancient - groups as the austral frogs, side-necked turtles, ratite birds and parrots, and the monotremes and marsupials. A common ancestry could now be entertained for their vicariant stocks isolated on the southern continents. These appeared to form a thin faunal base in Australia, over which a dominant immigrant Eurasian facies had later been imprinted. Secondly, the position of Australia so far south at the beginning of supposed Eurasian vertebrate invasions led the proponents of invasion to speculate that the Asian stocks had arrived later and had radiated faster than had been thought. Australia, it was realised, could hardly have been a biotic vacuum for such a long time in its early history; it must have had a Gondwanan fauna. The apparent dearth of such a fauna, particularly in birds and reptiles, was explained away by the ghost of competition past on a continental scale: all but a few of the Gondwanan animals had been out-competed and obliterated by the immigrant Eurasian stocks. Vertebrate zoogeography was still holding onto dispersalistic theory to explain the origin of the Australasian fauna. Indeed, it pervades Australian biogeographic thinking into the 1980's. Pick up any modern text and you will see bird geographers and reptile geographers talking about antarctic dispersal routes into Australia via Gondwana and Indo-Malayan dispersal routes in via Indonesia. Even current phytogeographic treatises talk about Australia receiving its first stocks of angiosperms by north-west land bridges from Laurasia in the Cretaceous. The point I want to make here, and I can't stress it enough, is that whatever biotic elements Australia received before its break from Antarctica in the early Tertiary it inherited from Gondwana. If angiosperms did come into the region from the north in the Cretaceous, they came to Gondwana, perhaps even the Australian-sector of Gondwana; but not to Australia as such. This point needs absorbing in Australian biogeographic thinking. The concept of dispersal routes, as it is being used today in Australasian biogeography, is all too often misused, confusing the difference between vicariance and dispersal at continental level, and the different contributions that both - and I stress both - have made to shaping the distribution patterns of plants and animals in the world today. So much for presumptions, what of the evidence? Enter the Australasian fossil record. For plants it is moderately good through the Tertiary, the critical period for the radiation of the flora; and, thanks to the palaeopalynologists, moderately well-worked over the last 20-30 years. It reveals that from the late Cretaceous to some 20-18 million years ago in the Miocene, Australia, New Zealand and what there was of New Guinea were clothed in temperate to subtropical rainforests little different in floristic composition from the temperate and subtropical rainforests of east Australia, montane New Guinea and New Zealand today. Nothofagus forests of tropical N. brassii and temperate N. fusca were widespread and so too the podocarps, laurels and myrtles. These forests today hold many primitive angiosperms - the Winteraceae, ancestral Proteaceae, and all the palaeopolypioid families of that great order Laurales - and they range from montane New Guinea and New Caledonia through east Australia and New Zealand to Chile in South America. According to traditional phytogeographic theory, these forests made up much of the newer immigrant Antarctic and Indo-Malayan elements in the Australasian flora. Where was the supposedly old autochthonous scleromorphic element? According to the fossil record, Eucalyptus, one of its most characteristic components, does not appear until Oligocene times, some 35-40 million years ago. So, as Bryan Badow has recently pointed out, the original pan-Australian phanerogam flora from the Cretaceous to mid Tertiary times - the flora that would have fed and sheltered the original stocks of Australasian vertebrates - was a Gondwanan, not autochthonous scleromorphic one. The scleromorphic flora came later. The fossil record for the fauna is much patchier and more poorly worked. In New Guinea found deposits only go back to the upper Tertiary 6 million years ago. In New Zealand, there are random early Tertiary deposits of marine vertebrates, then only one massive 'window' in the Quaternary, within the last million or so years. It comprises members of today's fauna plus the Moas. Australia is better served with two such 'windows on the past', one in the Pleistocene and the other, most fortuitously, in the middle Tertiary, from the late Oligocene to mid Miocene. Both windows have been worked actively over the last 20 years, work that is continuing today. The Pleistocene window reveals a massive fauna, larger and more diverse than todays, but still comprising the same groups of vertebrates - side-necked tortoises, giant goannas, diprotodonts, marsupial lions, giant kangaroos, a great radiation of ratite birds, giant mound-building fowls, eagles, and a rich flamingo fauna in the Lake Eyre basin. Flamingos today are centred in the southern continents of Africa and South America. These mega-elements of Australia's fauna were finally exterminated only about 15-18,000 years ago, at what seems to have been the combined hands of the last and glacial epoch and aboriginal man. The mid Tertiary window is even more informative. It comprises, once more, the same basic elements as Australia's vertebrate fauna does today, except they are more primitive in form. There are even platypusses with teeth; but there are no Eurasian rats or bats. The passerine or songbird component is still more interesting and is being worked out by Walter Boles at the Australian Museum. It includes lyrebirds and log-runners, one of which is virtually undistinguishable from a species found today in both montane New Guinea and central east coastal Australia. These birds are inhabitants of cool subtropical rainforests of the type dominated by Antarctic beech, laurels and myrtles; and their fossils are the oldest of any songbird yet found in the world. While the fossil record may provide good clues to the origin and radiation of Australasia's biota, it can never, on its own, provide final answers. It is too patchy stratigraphically, too limited geographically and too biassed against the less easily preserved organisms. The connections indicated by the fossil record between the Gondwanan rainforests and their faunas in montane New Guinea and Australia had, however, already been picked up in analyses of the Australasian bird and mammals faunas by John Calaby and myself in the early 1970's. We found that the birds and mammals of the montane New Guinean rainforests had their closest affinities not with those in the tropical rainforests of lowland New Guinea and Cape York Peninsula, but with others further south in Australia, from the Atherton Tablelands southwards. This was not a pattern of immigrant dispersal of indo-Malayan elements into Australia, but one of vicariance. And so we interpreted the faunas of these forests as old, relictual and ancestral to the bird and mammal faunas that had radiated in Australia's scleromorphic flora. But if current palaeontological and geographical evidence seem to be bringing together a picture of a prevailingly Gondwanan origin for the Australasian biota, they have so far failed to resolve at least two critical anomalies. One of these is in birds. If the Australasian bird fauna arose in Gondwana one might expect it to have close ties with the South American. Apart from a few groups of non-passerines, it doesn't; its passerines, in particular, do not. Passerines or songbirds are the most prolific order of birds on earth. Over half the world's birds are passerines, and they comprise over half the bird species in Australasia and South America. They are distinguished from other birds by free plantar tendons in the feet, which allows them to grip perches involuntarily, and by the structure and musculature of their syringes or voice boxes, which allows them to sing and defend territories by throwing their voices about instead of fighting. This has contributed to their success. South America's songbird fauna is dominated by a raft of endemic and sub-endemic families with unspecialised voice boxes. Australia's, by contrast, is dominated by families with conventional voice boxes and which otherwise resemble the songbirds of Afro-Asia. In the absence of a complete fossil record, finer tuning of these anomalies in birds is only possible through a finer tuning of phylogenetic links, something that comparative morphology has not been able to achieve after 200 years of research in this the most studied group of organisms in the world. New sets of characters were needed. Enter molecular genetics and the U.S. ornithologist Chades G. Sibley. No doubt you have all heard at this meeting of how molecular genetics has become the new whizzbang technology for systematic biology. And I expect that buzz acronyms like PCR, RFLP, ribosomal DNA, DNA fingerprinting, and DNA hybridisation have been echoing in the corridors. Today there is a wide and growing range of molecular and biochemical techniques - all good and with different values and virtues - for attacking different kinds of taxonomic and phylogenetic problems. When Sibley began assaying blood proteins in birds in the 1950's, however, there was little else. He didn't get far with blood proteins, then switched to egg-white proteins in the 1960's, and at the IOC in Canberra in 1974 presented results which suggested that Australia's divergent families of songbirds were all more closely related to one another than to their look-alikes in Eurasia. Ernst Mayr likened the results to reading tea-leaves in a cup, and so everyone forgot about them. Sibley went home to be fined for holding birds eggs illegally and switched attention to DNA-DNA hybridisation for salvation. DNA-DNA hybridisation is a black box/cook book technology in which single stranded DNA of one organism is bound with single copy strands of another and then heated until they dissociate. The temperature of dissociation gives a measure of the genetic relatedness between the two organisms across the entire nuclear genome. Sibley published his DNA hybridisation phylogenies for the world's songbirds in the mid 1980's. They indicated three primary assemblages. One was the morphologically primitive group centred in South America. Another was the honeyeater-scrub wren assemblage of Australia and New Guinea, linked to which were cuckoo-shrikes, birds-of-paradise, robins, monarch flycatchers and crow-shrikes also centred in the same region. And last was the thrush-warbler-lark-finch assemblage of Eurasia and North America. It was a sensational result. Overnight, the Australo-Papuan songbirds found themselves landed with a marsupial-like radiation. Ernst Mayr eulogised that Sibley had scaled the ice wall. Had Sibley made it at last? Oh no. He turned his attention to the ape and man, stirred up a predictable hornet's nest, and, if you've been following it in Science, you'll know he's under fire for using an unsatisfactory constant in his measures of dissociation and for discarding data points that didn't fit as they should. It is all too easy to forget, through this flack, that Sibley has been the greatest pioneer yet in the use of biotechnology for systematic biology. Sibleys results in songbirds needed testing, either by re-runs of DNA-DNA hybridisation or, preferably, with other, independent molecular and biochemical evidence. This is now coming from work in which I am collaborating with Dr Peter Baverstock of the Northern Rivers College of Advanced Education and with Dr Les Christidis at the Museum of Victoria. Baverstock is using an immunological technique called microcomplement fixation and Christidis protein electrophoresis. The first is a rather approximate black box technology; and the second is limited by the number of characters or proteins that can be screened, abetted by the possibility of convergence in their alielic states at the level of familial comparisons. Whatever the shortcomings of these techniques, however, their results, except on several minor points, are broadly consistent with the radiations indicated by DNA-DNA hybridisation. The diverse Australo-Papuan centred songbird families, although closer as a group to Eurasian than South American songbirds, were nevertheless allied still more closely to one another. Thus the Australasian flycatchers are not related to Eurasian flycatchers but to Australian butcherbirds and cuckoo-shrikes; and the Australian wrens and warblers are not related to their namesakes in Eurasia, but to the Australasian honeyeaters and robins. These results reveal the similarities in form between Australasian and Eurasian songbirds as superficial convergences that have arisen in adaptation to the same sorts of niches and the same ways of life. More than that, and this is important, the electrophoretic data indicate that the genetic distances between the Australasian songbird families are greater than those between the Eurasian. If genetic distance is time-related, as I believe it is even if we don't yet have an accurately calibrated molecular clock, then this means that the Australasian songbird families are older than those in Eurasia. Think of its implications. If the Australasian and Eurasian songbird groups are monophyletic - and there is considerable morphological and molecular evidence indicating that they are - then the Eurasian groups are derived from the Australian, in reverse to the traditional view. Carry this concept on to the supposedly Eurasian reptiles centred in Australia - the diplodactyline geckos, goannas and dragons - and suddenly the entire Australasian land vertebrate fauna begins to take on a prevailingly Gondwanan aspect. In this scenario, the world's songbirds are Gondwanan and on that supercontinent diverged into eastern and western groups; the western group, primitive with an unspecialised voice box, was inherited by South America; and the eastern group with a specialised voice box was inherited by Australia. From there it colonised Eurasia later. From all this data - old morphological and new palaeontological and molecular - a broad new picture emerges of the origin and radiation of Australasia's phanerogam flora and vertebrate fauna. Australia and New Zealand inherited from Gondwana a prevailing cool temperate to subtropical rainforest biota, one dominated by podocarps, antarctic beeches, laurels, myrtles, proteads, cunoniads, elaeocarps and Winteraceae among plants, and dare I add, monimiads and even pittosporums. The vertebrate fauna included the primitive myobatrachid and leiopelmatid frogs, sidenecked land turtles, the range of land reptiles now centred in both lands, plus ratite birds, parrots, many other non-passerines, and the ancestral specialised songbirds. Breaking away first, New Zealand may have missed out on monotremes and marsupials; and being small and at times broken, it had much less space for diversification than Australia. Its biota was probably never particularly rich, but it probably also changed rather little under a moderately constant climate on its northward drift through the Tertiary. Man and the Plio-Pleistocene glacials brought moderate extinction in recent times but proximity to Australia then allowed wind-blown colonisation by a range of Australian bird stocks and several bats. Australia itself had also inherited a rather diverse waterbird fauna, including flamingos, on lakes across its central Artesian basin. From the later Miocene on, however, climate became progressively drier, beginning about 17-18 million years ago. The aquatic fauna dwindled and the rainforest biotas retreated to the wetter periphery of the continent. This gave the impetus for the radiation of the scleromorphic flora, the ancestral elements of which had evidently developed endemically from hardier rainforest stocks and now diversified to replace the retreating rainforest. Elements of the land fauna radiated with it, derived from ancestral Gondwanan rainforest stocks. Climatic deterioration peaked in the dry and cold glacial oscillations of the Plio-Pleistocene, when, fortuitously, the present Great Dividing Range and central cordilleras of New Guinea reached their greatest rain-catching height. New Guinea by this time had reached its present form and fronted broadly along the northern edge of Australia; its last connection was severed only 11,000 years ago by the flooding of Torres Strait. The Tertiary rainforest biotas completed their retreat to refugial pockets in the Great Dividing Range and to subtropical and temperate levels in montane New Guinea, in reverse to the traditional view of their advent in Australia. As Michael Archer has pointed out in his compendium of "Vertebrate Zoogeography and Evolution in Australia", a walk up a New Guinea mountain is like a walk back through time. In the New Guinea mountains, the Australian biota became intermingled with the Malesian rainforest flora, a flora which completely dominates the lower altitudes on that island and on east Cape York Peninsula. Curiously, the vertebrate fauna of the Malesian forests in New Guinea is almost completely Australian; it is the other significant anomaly that I referred to earlier and it has yet to be explained. The coincidence of increasing aridity with the approach of Australia and its fringing islands to Asia at the end of the Tertiary developed Australia's arid zone biota and made space for Eurasian immigrants. The exotic nature of Australia's chenopod flora and northern Australian savannahs is well documented; and they have provided niches for radiation not so much in Australian bird families as in Eurasian sylviid warblers. Except for reptiles, the arid zone biota, covering some 70% of Australia today, is the poorest of all in vertebrates. If the old rainforest blota was the reservoir for adaptive colonisation of the and zone, and the scleromorphic biota the plumbing and taps to feed it, the arid zone has behaved like a sink, its plug pulled more-or-less by every dry glacial cycle during the Plio-Pleistocene. These scenarios can be encapsulated by minor but significant modifications to the traditional concepts of Australian biotic elements. For the broad patterns of distribution have not changed, even if the interpretation of their origin has. All that I have had to do here to conventional schemes of southern Bassian, northern Torresian and central Eyrean or Eremaean biotas is to separate the Malesian or Irian rainforest biota and the old Australian Tertiary or Tumbunan rainforest biota out from the Torresian eucalypt element, and, on top of that, to superimpose arrowed lines showing the directions of adaptive radiation (Fig. 1). The word Tumbunan, incidentally, comes from the Melanesian pidgin word Tumbuna, meaning ancestor.
After all this, do you have a sense of deja vu? In the middle of this century, plant and animal geographers presented a combined picture of the origin and radiation of Australia's higher order biota with all the hallmarks of finality. Now, only a few decades later, we are proposing another, almost diametrically opposed view with a similar certainty. This carries a three point cautionary message, of which I know Nancy Burbidge would have approved: First, biogeographic reconstructions such as we have examined today are no more or less than best-fit hypotheses for the data available. They stand only by continuing testing. Secondly, as many sets of evidence as possible should be enjoined to resolve biogeographies, not just present distribution patterns or the fossil record but phylogeny, biotechnology and evolutionary ecology as well, and anything else that might help. And let us not forget the service that morphology has and will still render us. Thirdly and finally, a sound biogeography can only come from a sound taxonomy. Without it, biogeographic constructions, like other endeavours in biology extending to conservation ecology and the breeding of economic crops, are built on foundations of clay and will eventually crumble. top |