
Mosquito Orchids
Deciduous terrestrial orchids that reproduce from seed and also grow in clonal colonies, often with a low proportion of flowering plants. Leaf single, heart-shaped, usually reddish or purplish beneath. Leaf blade held horizontally above the soil surface on a slender stalk. Inflorescence a multiflowered raceme arising in the leaf base. Flowers small, right-way-up, insect-like, pale coloured. Sepals short, with short cylindrical or terete tips. Petals small, reflexed or spreading. Labellum proportionately large, fixed to the base of the column. Labellum lamina projecting forwards, concave at the base, with short wart-like papillae. Callus with a pair of prostrate comma-shaped basal glands and a central channel. Column long, slender, not winged, arching over the labellum base.
Similar Genera
Significant Generic Characters
Deciduous terrestrial; leaf single, about as long as wide, usually reddish beneath; raceme multiflowered; flowers small, pale-coloured, resupinate; sepals with short terete osmophores; dorsal sepal larger than the lateral sepals; petals small; labellum unlobed, base concave, margins denticulate; callus dominating the labellum lamina, papillate, with 2 prostrate glands and a central channel; column not winged.
Size and Distribution
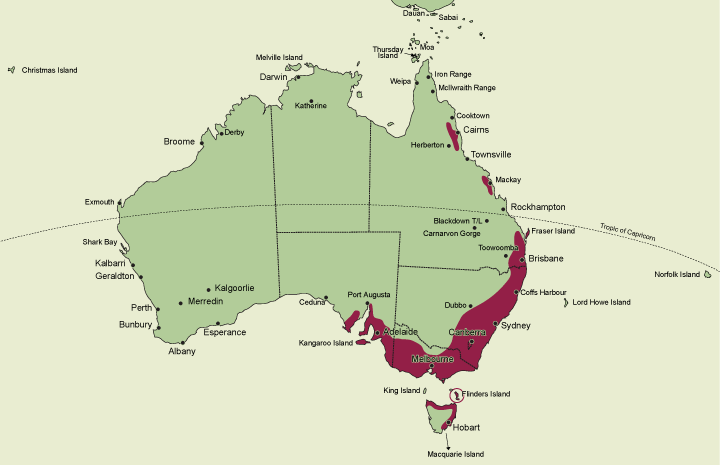
A genus of 8 species of terrestrial orchid, 7 species occurring in eastern Australia, (including Tasmania) and a single species in New Zealand. In Australia the genus is distributed from the Windsor Tableland in northeastern Queensland (17�20' S) to the southern tip of Tasmania (43� S) and west to Eyre Peninsula in South Australia (135�30' E). They extend from coastal areas to inland ranges but are absent from semi-arid inland areas and subalpine and alpine zones. State occurrence: Queensland, New South Wales, Victoria, Tasmania, South Australia.
Ecology
Acianthus plants grow on sheltered slopes, near streams, in gullies and under shrubs and bracken in grassy to shrubby sclerophyll forest, woodland, coastal scrub and heathland. Most species favour shady situations where the humidity is high during the growing season, but with free and unimpeded air movement. Soils are well-drained sands, sandy loams and clay loams.
Biology
Pollination: The flowers of the Australian Acianthus species open in succession up the raceme, with each flower lasting several days. Pollination of 7 of the Australian species of Acianthus is by small flies of the families Anisopodidae, Sciaridae and Mycetophilidae (Cheeseman 1875, Jones 1974, Bower 2001). These flies feed on nectar produced by the basal labellum glands and which gathers in droplets in a sunken area of the labellum base. By contrast the flowers of Acianthus exiguus are self-pollinating. Apomixis is unknown in the genus.
Reproduction: All species of Acianthus form dense, sometimes extensive, clonal colonies which extend annually by means of daughter tubers produced freely on the end of long stolonoid roots. After pollination the capsules swell in an erect position and seed dispersal occurs 8-10 weeks later.
Seasonal Growth: These orchids occur in areas with a seasonal climate. Acianthus plants grow actively in autumn and winter and are dormant during the extremes of heat or dryness over summer, surviving these times as small fleshy tubers.
Flowering: Species of Acianthus flower between February and August, either producing the inflorescences as the leaves develop (Acianthus apprimus) or well after they have matured (Acianthus fornicatus). It is a usual occurrence that only a very low proportion of flowering plants will occur in a colony. Adverse climatic conditions in the early stages of inflorescence development frequently lead to the inflorescence aborting without maturing its flowers.
Hybrids: Natural hybridisation is very rare within Acianthus. Occasional hybrids have been collected between Acianthus fornicatus and Acianthus exsertus.
Fire: Growth and flowering of these orchids is inhibited for one or two seasons after hot summer fires.
Derivation
Acianthus is derived from the Greek acis, point, anthos, flower and refers to the slender points on the sepals of some species.
Botanical Description
Perennial geophytic herbs, sympodial. Plants glabrous except for multiseriate hairs on the stem and collar. Roots absent. Tubers globose, paired, fleshy; replacement tubers formed at the end of short droppers; daughter tubers formed at the apex of horizontal lateral stolonoid roots. Stolonoid roots and subterranean axes with minute papillae each with apical unicellular hairs. Stem erect, short, unbranched, terete, with membranous cataphylls at each node. Leaf lasting a single season, solitary, basal; lamina firm-textured, held horizontally above the soil surface, flat, hypostomatic, convolute in bud, sometimes lobed, usually with abaxial anthocyanin pigments. Venation anastomosing, the main veins uniting apically; petiole fused with base of scape. Inflorescence racemose, one-several-flowered, erect, terminal; scape lacking sterile bracts. Floral bracts foliaceous, small. Ovary elongate, ribbed, glabrous. Flowers resupinate, small, insectiform, dull coloured (pinkish to purplish), pedicellate. Nectar present. Dorsal sepal free, much broader than lateral sepals, with a short terete apical extension. Lateral sepals free, much narrower than the dorsal sepal, with a short terete apical extension. Petals free, much smaller than the sepals. Labellum free, porrect, attached by its base to the anterior column base, markedly dissimilar in size and shape to the sepals and petals, ecalcarate. Labellum lamina membranaceous, not lobed, concave; margins flat or revolute, denticulate; apex apiculate to acuminate. Spur absent. Callus fleshy, dominating most of the lamina, with two basal glands and a narrow reticulate longitudinal channel; ventral surface with subconical papillae. Basal glands prostrate, tonsil-like or comma-shaped, partially embracing the anterior base of the column, incurved distally, their apices weakly confluent. Column lacking free filament and style, slender; base enlarged on anterior side; apex dilated, incurved, gibbous behind anther. Column wings absent. Column foot absent. Pseudospur absent. Anther terminal, 2-celled, persistent, basifixed, porrect, not rostrate. Pollinarium present, consisting of 4 pollinia in 2 pairs, each pair attached to a small hemiviscidium. Pollinia deeply lobed, each lobe clavate, narrowed to a basal stalk, waxy, yellow; pollen grains in tetrads. Viscidia lateral to apex of rostellum. Rostellum ventral, 2-lobed. Stigma entire, small, concave. Capsules dehiscent, thin-walled, glabrous, erect; pedicels not elongating in fruit; peduncle not elongating in fruit. Seeds numerous, light coloured, oblong-ellipsoid, winged; cells aligned in rows.
Taxonomy
A revisionary treatment of Acianthus has been published (Kores 1995) but in this study the author adopted a very broad generic approach, for example including Cyrtostylis within the generic boundaries of Acianthus despite an earlier study to the contrary (Jones and Clements 1987). Kores recognised infrageneric taxa within his broad concept of Acianthus and did not recognise several segregate species described by Jones (1991). In more recent studies which affected Australian taxa, Szlachetko (2001) elevated Acianthus section Univiscidiatus to generic rank and Jones et al. (2002) defined Acianthus in the strict sense and erected Acianthella and the monotypic genus Nemacianthus (Jones and Clements 2004).
Notes
The leaves of all the Australian species of Acianthus are very similar and difficult to identify to species level without the flowers. Leaf expansion continues after emergence and the leaves expand considerably in size after flowering.
Nomenclature
Acianthus R.Br., Prodr. 321 (1810). Type species: Acianthus exsertus R. Br.
Infrageneric Taxa: No infrageneric taxa are recognised.
References
Bower, C.C. (2001). Pollination (Acianthus, pp. 69-70), in Pridgeon, A.M., Cribb, P.J., Chase, M.W. and Rasmussen, F.N. (eds), Genera Orchidacearum, vol. 2, Orchidoideae (part one). Oxford University Press, United Kingdom.
Cheeseman. T.F. (1875). On the fertilisation of Acianthus and Cyrtostylis. Transactions of the New Zealand Institute 7: 349-352.
Jones, D.L. (1991). New taxa of Australian Orchidaceae. Austral. Orch. Res. 1-207.
Jones, D.L., Clements, M.A., Sharma, I.K., Mackenzie, A.M. and Molloy, B.P.J. (2002). Nomenclatural notes arising from studies into the tribe Diurideae (Orchidaceae). Orchadian 13(10): 437-468.
Jones, D.L. and Clements, M.A. (1987). Reinstatement of the genus Cyrtostylis R. Br. and its relationships with Acianthus R. Br. (Orchidaceae). Lindleyana 2: 156-160.
Jones, D.L. and Clements, M.A. (2004). Acianthella, a new genus in the Acianthus alliance (Orchidaceae). Orchadian 14(7): 330-332.
Kores, P.J. (1995). A systematic study of the genus Acianthus (Orchidaceae: Diuridae). Allertonia 7(3): 87-220.
Szlachetko, D.L. (2001). Genera et Species Orchidalium 1. Polish Bot. J. 46(1): 11-26.